
Etiquetas
CMH, Gorer, Histocompatibilidad, HLA, MHC, Snell, Transplantes
| COMPLEJO HLA | |||||||
| Clase CMH | CMH-II | CMH-III | CMH-I | ||||
| Región | DP | DQ | DR | C4,C2,BF, | B | C | A |
| Productos genéricos | DP(a,b) | DQ(a,b) | DR (a,b) | proteinas del complemento, TNFa, TNFb | HLA-B | HLA-C | HLA-A |
| Ubicación | Hacia centromero | brazo corto cromosoma 6 | Hacia telómero |
En la homeostasis del hombre con el medio externo e interno, a lo largo de la evolución el Sistema de Histocompatibilidad ha ido adaptando al hombre a los alérgenos, dando respuesta adecuada contra los patógenos.
El complejo principal de histocompatibilidad (CMH), aparece en todas las especies de vertebrados y tiene un papel fundamental en la presentación a las células del sistema inmunológico.
La estructura del CMH se conoce en siete especies de mamíferos euterios (placentarios), dos de aves, cinco peces teleósteos y en los tiburones.
En los humanos, los 3,6 Mbp de la región MHC, del cromosoma 6, con 140 genes con los marcadores genéticos MOG Y COL HA2.
En los años 30 de este siglo, Gorer&Snell (premio nobel en 1980) , estudiando los antígenos de superficie de células sanguíneas, identificaron varios grupos de genes responsables de los antígenos. Los cuales estaban estrechamente ligados, determinando el rechazo de transplantes entre individuos no emparentados de la misma especie, a estas moléculas se las denomino antígenos de histocompatibilidad y al conjunto de genes ligados que los codifican complejo mayor de histocompatibilidad. En los seres humanos el CMH, llamado HLA (human leukocyte antigen) se descubrió a través del análisis de transfusiones sanguíneas y transplantes de órganos.
En cada especie de mamíferos los distintos loci del CMH son muy polimórficos, poseen la mayor variabilidad intraespecífica detectada en la Genética de Poblaciones, cada locus dentro del CMH posee multitud de variantes alélicas dentro de poblaciones naturales de cada especie. Cada individuo hereda un juego de CMH del padre y otro juego de la madre, cada uno de los juegos completos heredados de un progenitor se denomina haplotipo. Los dos alelos de cada locus son de expresión codominante: un individuo heterozigoto para los distintos loci del CMH expresa en sus células al mismo tiempo los dos tipos de variantes alélicas de cada locus.
El polimorfismo de cada locus dentro de poblaciones normales hace que las poblaciones resistan el ataque de gran variedad de patógenos, aunque algunos individuos resulten poco aptos. La importancia adaptativa del polimorfismo CMH en una población es que tiende a proteger a la especie frente a agentes infecciosos, ampliando la variedad de antígenos que se pueden reconocer.
Cuando disminuye el grado de polimorfismo del CMH, aumenta el riesgo de enfermedades infecciosas en las poblaciones. Para generarse en las poblaciones de vertebrados este polimorfismo:
- recombinación homóloga entre alelos del mismo locus
- conversión génica: una secuencia de un alelo de un locus del CMH es reemplazada por otra secuencia de un gen homólogo.
- mutaciones puntuales
Las moléculas del CMH muestran una gran especificidad para unirse a los péptidos, el peptido que se une presenta una serie de características: una es el tamaño, también secuencias de aminoácidos que permitan complementariedad, y para ser capaz de activar el linfocito T poder encajar en la hendidura de la molécula del CMH. La velocidad de asociación es muy baja, pero la velocidad de disociación es aun más baja para interaccionar con el linfocito T, las asociaciones de péptidos a las moléculas del CMH son saturables y de baja afinidad. Otra característica es que se pueden presentar antígenos exógenos como propios, permitiendo solo la supervivencia de los linfocitos T que no reaccionan contra el organismo.
Todas las moléculas del CMH poseen 4 segmentos. Un segmento de unión al péptido o hendidura, un dominio tipo Inmunoglobulina (Ig), un segmento transmembrana y una porción citoplasmática carboxi-terminal.

Las moléculas de clase I del CMH se expresan en todas las células nucleadas, es un mecanismo de defensa efectivo en células nucleadas infectadas, al no poder migrar, en la única manera de que los linfocitos T CD8+ siendo presentados los péptidos por el CMH I lisen las células infectadas.
Presentan antígenos endógenos.
Las moléculas de clase II del CMH se expresan en celulas presentadoras de antígeno: linfocitos B, Macrófagos y células Dendríticas. Estas células reconocen, fagocitan, procesan y presentan en su superficie celular a los péptidos exógenos unidos a las moléculas de clase II.
Presentan antígenos exógenos.
Las moléculas tanto de clase I como de clase II pueden AUMENTAR LA EXPRESIÓN, afectadas por las citoquinas secretadas en la inmunidad innata como en la inmunidad adaptativa.
- INFalfa, beta: respuesta inmunitaria frente a los virus.
- TNF(factor de necrosis tumoral), LT (linfotoxinas): se liberan en las infecciones microbianas.
La inmunidad innata estimula a la inmunidad adaptativa.
- los linfocitos B aumentan su expresión bajo IL-4.
- las células dendríticas aumentan su expresión a medida que maduran.
PROCESAMIENTO DE LOS ANTÍGENOS
Los peptidos exógenos son internalizados, procesados, expresados en la superficie celular unidos a moléculas de clase II del CMH y reconocidos por los Linfocitos T CD4+ , los peptidos endógenos son presentados unidos a moléculas de clase I a los Linfocitos T CD8+.
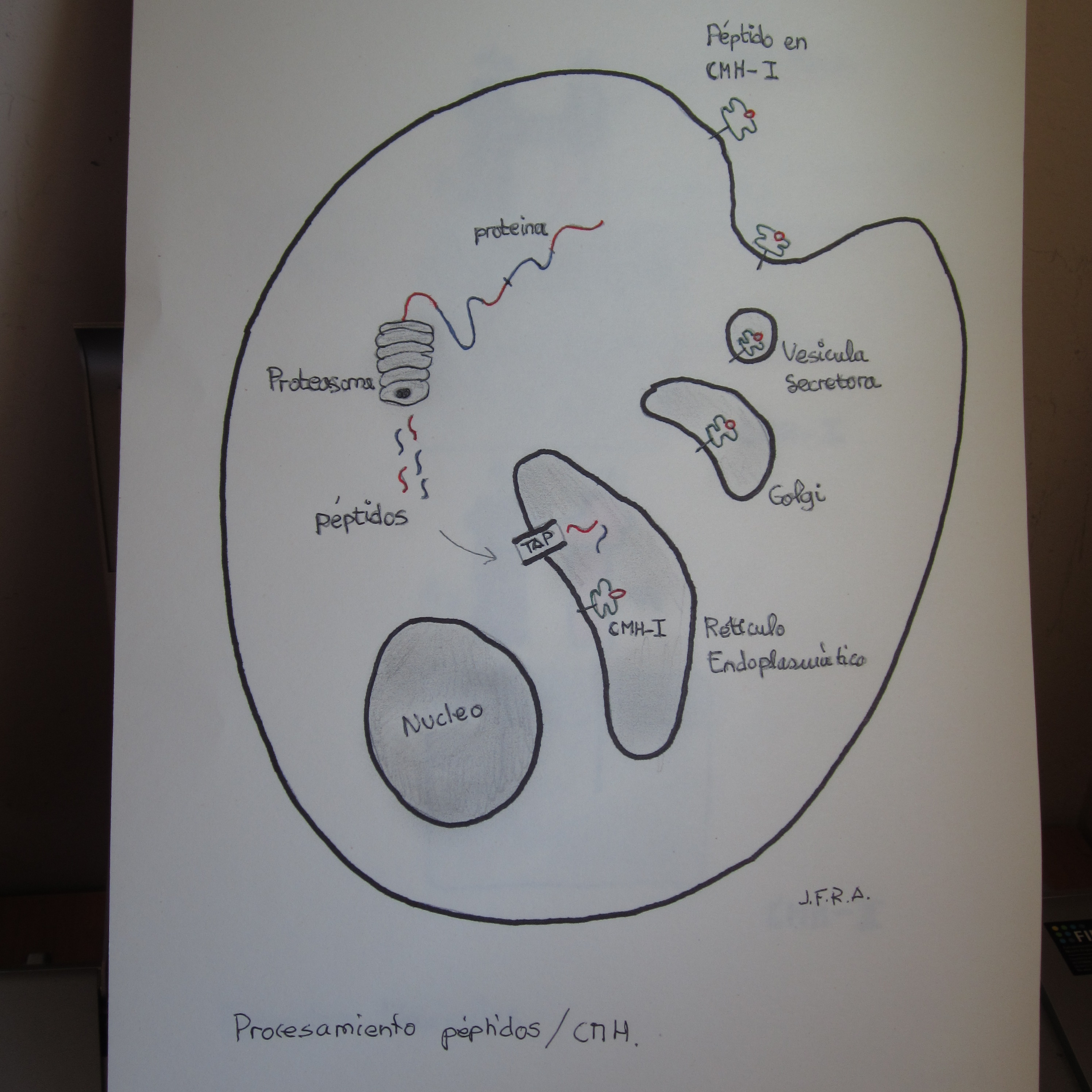
Veamos con un poco mas de detenimiento el procesamiento de péptidos asociados a moléculas CMH-1: proteinas presentes en el citosol son degradadas por el proteasoma, y los péptidos resultantes son internalizados por el canal TAP en el retículo endoplasmático, donde se asocian con las moléculas sintetizadas de CMH-1. Los complejos péptido-CMH-1 pasan al Aparato de Golgi, donde son glucosilados, y de ahí a vesículas secretoras, que se fusinan con la membrana celular, de forma que los complejos quedan expuestos hacia el exterior, permitiendo el contacto con los linfocitos T circulantes.
INMUNOLOGÍA Y TRANSPLANTES
El transplante consiste en tomar células, tejidos u órganos, llamados injertos de un individuo y ponerlos en otro individuo. El individuo que proporciona el tejido es el donante y el que lo recibe receptor o huésped.
Los injertos pueden ser del mismo individuo, o de otro de la misma especie o de otras especies. El primero es injerto autógeno, a los individuos de la misma especie se los denomina injerto alogénico y aquellos de distinta especie injerto xenogénico.
El CMH reconoce el injerto extraño de dos maneras:
- presentación directa: es exclusiva de las moléculas del CMH extraño, implica el reconocimiento de una moleculas del CMH intacta ofrecida por APC del donante existentes en el injerto y se debe a la similitud entre la estructura de la molécula del CMH extraño (alomolécula) intacta y las moléculas del CMH propias.
- Presentación indirecta: supone el procesamiento de las moléculas del CMH del donante por parte de las APC de receptor y de la presentación de los péptidos derivados de las alomoléculas del CMH asociadas a moléculas del CMH propio.
Los rechazos a los transplantes se clasifican:
- Hiperagudo: oclusión trombótica de la vasculatura del injerto, que comienza a minutos u horas de la anastomosis entre los vasos del donante y el huésped.
- Agudo: es mediado por las células T y B activas, que producen injuria parenquimatosa y vascular del injerto. Comienza a la semana del transplante, que es el tiempo que toma la inmunidad adaptativa en dar inicio a sus mecanismos efectores.
- Crónico: fibrosis y alteraciones vasculares, con perdida de la función del injerto durante un periodo prolongado, la fibrosis del rechazo crónica puede deberse a reacciones inmunitarias y a la síntesis de citoquinas que estimulan a los fibroblastos.
La estrategia de la práctica clínica ante el rechazo es inmunodepresión general, reducción de la intensidad de la aloreacción específica y la tolerancia específica al aloinjerto.
